スパインアクチン細胞骨格は興奮性シナプス成熟を制御する
関野祐子・高橋秀人・白尾智明
群馬大学・院・医学系研究科・高次細胞機能学分野
Department
of Neurobiology and Behavior,
英文タイトル:Actin cytoskeleton of the dendritic spine governs maturation of
excitatory synapses
キーワード:樹状突起フィロポディア,スパイン,アクチン,ドレブリン,PDS-95
要約
成熟した脳の錐体神経細胞の樹状突起には無数のスパインがあり,グルタミン酸受容体や,シナプス後部肥厚タンパク群,アクチン細胞骨格タンパク群が階層的に集積している。我々は最近,樹状突起フィロポディア内のドレブリン結合型アクチンの集積が、シナプス後部肥厚タンパクの集積を引き起こし、スパイン形成を制御することを見いだした。スパインアクチン細胞骨格の形成こそが、興奮性シナプス成熟の本質であると述べたい。
はじめに
脳は,遺伝子の情報に従って中枢神経細胞の数と配置を決定したあと、環境情報をもとに神経細胞同士の結合(シナプス)の数を増やしたり減らしたりしながら成熟する。この時にシナプスそのものも未熟な状態から成熟した状態へと変化する。中枢神経系の主な興奮性神経伝達物質であるグルタミン酸によるシナプスは,樹状突起上にある無数の小さな(約1 m)スパイン(棘状突起)に形成されている(Shepherd,
1996)。(Leuner et al., 2003)(図1−A;培養後21日ラット海馬神経細胞の樹状突起)。
スパインは、脳機能の発達・成熟に伴って特徴的な形態変化を示す。したがって,スパイン形成は脳機能成熟のメカニズムを解く鍵として強い関心を集めて来た。生まれたばかりのネズミの脳の神経細胞樹状突起には,スパインはまだ出来ていない。その代わり,フィロポディアという,スパインより細長い突起が多数存在する(図1−B;培養後7日ラット海馬神経細胞の樹状突起)。生後2週頃からスパインの数は急激に増加する(Harris
et al., 1992; Boyer et al., 1998)。いったいどのようにしてスパインが出来てくるのであろうか。フィロポディアがそのままスパインに変化するのであろうか。それともフィロポディアとは全く関係なく、伸びてきた軸索終末が樹状突起幹に接触して樹状突起幹シナプスができてから,それがスパインとして突出してくるのであろうか。スパインが初めて見出されてからすでに100年以上経つが,スパイン形成過程はその分子機構も含めて謎のままであった。
ところが最近,これまでの古典的研究とはまったく異なるストラテジーにより,この謎解きに解決の糸口が見えてきたのである。そのひとつは、それは,スパインシナプスを構成する分子群(細胞骨格タンパク群,シナプス後部肥厚を構成するタンパク群、カルシウム結合タンパクやリン酸化酵素などシグナル伝達分子群)が次々と明らかにされたことである。また,神経細胞培養技術や遺伝子工学技術が普及したことも挙げられる。本稿ではとりわけ,フィロポディアがスパインに変化する時の形態変化の分子機構について紹介する。
スパインがどのようにして出来上がるかを知るためには,完成したスパインの姿をまず知っておかなければならない。そこで,スパインの微細構造とその構成タンパクについて,概説する。
1.スパインの形
ゴルジ鍍銀法により観察される大脳皮質錐体細胞の典型的なスパインは,長さ1−2m,幅が0.5−1 mほどの大きさである。多くのスパインの先端は、丸みを帯びており、これをスパイン頭部という。スパイン頭部の大きさはシナプス前終末のシナプス小胞数と相関しており(Harris
and Stevens, 1989),スパイン内のカルシウム動態も個々のスパイン形態に依存している(Volfovsky et al., 1999)。また,AMPA型グルタミン酸受容体は頭部の大きなスパインに局在し(Takumi
et al., 1999; Matsuzaki et al., 2001),スパイン形態の安定性もスパインに分布するグルタミン酸受容体の種類に相関している。このように,スパインの形の違いには,カルシウム動態の違いやシナプス電流の種類や大きさの違いが反映している。
2.スパインの微細構造
大きさも形も反応性も様々であるスパインに共通する電子顕微鏡レベルの微細構造の特徴は,2つある。すなわち,シナプス後膜直下にシナプス後部肥厚(postsynaptic density;PSD )を持つことと,スパイン頭部はアクチン繊維が密集した細胞骨格で出来ていることである。
スパインには,シナプス後膜直下に高電子密度の構造体すなわちPSDとよばれる裏打ち構造を持つ非対称型のシナプス接合部分がある。スパインのシナプス後部膜には、イオン透過型グルタミン酸受容体と代謝型グルタミン酸受容体が密集している(Nusser et al., 1994)。一方,対称型を呈するシナプスは-アミノ酪酸やグリシン作働性の抑制性シナプスであり,樹状突起上,スパイン起始部,細胞体周辺部に見出される(Shepherd,
1996)。このことは,スパインは興奮性伝達物質であるグルタミン酸をうけとるために特殊化した構造であることを示している。
もう一つのスパインの特徴は,アクチンが高密度集積していることである(LeBeux and Willemot, 1975; Fifkova and Delay, 1982; Matus et al., 1982)。神経細胞の細胞骨格は微小管・ニューロフィラメント・アクチンフィラメントの3種類であるが,アクチンフィラメントは、とりわけスパインに高密度に集積している。おもしろいことに,樹状突起幹の細胞骨格である微小管はスパインには入り込んでいない(Fifkova, 1985)。このことから,スパインが樹状突起とは独立した構造であることがわかる。
PSDの細胞質側には,スパイン内部とは異なる編み込み状のアクチン繊維構造がある。そのためPSDは受容体シグナルをアクチン細胞骨格に伝えるトランスデューサーのような位置関係にあると考えられる。確かにPSDには,タンパク同士をつなぎ止める役割を果たすタンパク群があり,受容体とその他のタンパクとアクチン細胞骨格をつなぎ止めるアンカーの働きをしている。
つまり,スパインはグルタミン酸受容体を介した信号伝達を処理するために,PSDとアクチン細胞骨格による階層的な構造をしている。個々のスパインは一つの興奮性シナプスの情報を処理する最小単位構造であるといえる。
II
. スパインの構成タンパク
スパインの形成機構を明らかにするためには,PSDとスパインアクチン細胞骨格を作っているタンパクの動態やそれらの相互作用を明らかにすることが重要である。本稿では,PSDの代表的な構成タンパクPSD-95とスパインアクチン細胞骨格を作るアクチン結合タンパクドレブリンに着目する。
1.代表的なシナプス後部肥厚タンパク:PSD-95
PSD-95は、memmbrane-associated guanylate kinase
(MAGUK)と総称されるタンパクの一つである。PDZドメインとSH3ドメインに加えてグアニル酸キナーゼ(GK)ドメインという複数のタンパクータンパク相互結合部位を持つので,相互に多量体を形成しシナプス後膜直下で2次元の格子を形成しているモデルが考えられている(Sheng
and Sala, 2001)。PSD-95は、NMDA型グルタミン酸受容体NR2サブユニットと他のタンパクを集合させてシグナリングコンプレックスを形成している(Kennedy,
1997; Ziff, 1997; Sheng and Sala, 2001)。AMPA型受容体はすべてのスパインに存在するのではなく,頭部の大きなスパインに局在しるのに対して,NMDA受容体はサイレントシナプスも含めてどのスパインにも存在する。従って、NMDA受容体のアンカリングタンパクであるPSD-95は,スパインのPSD構造を検出するための有用な組織化学マーカーであるといえる。
2.スパインのアクチン結合タンパク:ドレブリン
ドレブリンは,脳の発達に伴って発現量変化やアイソフォーム変換がおこるタンパクとして同定された(Kojima
et al., 1988; Shirao et al., 1988)もので,中枢神経系で見出されたアクチン結合タンパクの中で最も発現量が多いものの一つで,神経系の代表的なアクチン結合タンパクであるといえる。成熟した神経細胞ではドレブリンAアイソフォームが主体で,ラットの場合では生後1週間以降に急速に発現が増加して,未熟神経細胞型ドレブリンEアイソフォームの発現は減少する。ドレブリンAの発現急増がシナプス形成の時期と一致すること,ドレブリンが成熟した神経細胞のスパインに局在すること(Hayashi
et al., 1996)などは,ドレブリンAとスパイン形成との間の深いつながりを想像させる。ドレブリンは,トロポミオシンを競合的に追い出してアクチン繊維に結合する。このときアクチン繊維は,比較的短くて太いアクチン繊維の束がカーブしたり丸くなったりしたような状態になる(Shirao
et al., 1994)。ドレブリンが発現した線維芽細胞は,もはや扁平の形をしなくなり,細胞体は丸く,細長い神経細胞様突起を形成するようになる。トロポミオシンのついたアクチン繊維の束にはゲルゾリンが入り込めないのでアクチン繊維の断裂はおきにくいが,ドレブリンがついているアクチン繊維にはゲルゾリンが入り込めるようになり,アクチン繊維はある程度の長さで断裂されるようになる。また,
ドレブリンはアクチン−ミオシン相互作用を抑制するので,一度できた突起は退縮しにくくなる。またドレブリンは,-アクチニンとアクチン繊維の結合を競合的に拮抗するので,-アクチニンと結合するNMDA型受容体の活性調節に関与している可能性がある。このような生化学的性質から,スパインのアクチン細胞骨格は、ドレブリン結合型アクチンにより特徴付けられているといえる。
我々がドレブリンに着目した理由のひとつは、アクチン細胞骨格を論じる際に、アクチン結合タンパクの存在を常に考慮しなくてはならないからである。建築物にたとえれば,アクチン繊維そのものは鉄骨であり,それ単独では形を作る骨格にはなり得ないからである。アクチン繊維の長さや曲がり易さや形状は,アクチン結合タンパクにより決定されている。たとえば、アクチン繊維にトロポミオシンが結合するとストレスファイバーとなり,線維芽細胞などが扁平な形態となる。また,軸索が伸展するときには,その先端の成長円錐のさらに先端の細い突起の中では,アクチン繊維はファシンによって束ねられている。このように,アクチン繊維は細胞内の部位特異的なアクチン結合タンパクと結合して,部位特異的なアクチン細胞骨格を作る。大げさにいえば,アクチン結合タンパクが、その部位のアクチン細胞骨格の性質と機能を決定していることになる。その点で、ドレブリンはスパイン内のアクチンフィラメントに結合し、スパイン特異的なアクチン細胞骨格の形成を担っているといえよう。
III.スパイン形成のシグナリングの解明
さて,スパインの主要な構造とその構成タンパクの代表として,PSD-95とドレブリンを取り上げたが,これらのタンパクがいったいいつ,どこに,どのような時間経過で,集積するのであろうか。そして,それらの集積は全く独立に起こるのか,それとも相互に依存して起こるのであろうか。
1.フィロポディアはスパイン前駆体であるのか。
スパインの出来方には,大きく分けて3つのモデルがある。Harrisらは,樹状突起フィロポディアが軸索に接触してから退縮し,軸索を樹状突起に引き込んでひとたび樹状突起幹シナプスとなり,そこからスパインが成熟するというモデルを提唱した(Fiala
et al., 1998; Harris, 1999)。Craigらは,未熟な神経細胞では樹状突起幹シナプスにPSD-95クラスターが見られることから,軸索が樹状突起に接触してシナプスを作り,そこからスパインが生ずるというモデルを提唱した(Rao et al., 1998; Rao and Craig,
2000)。これらのモデルに対して,Smithらは,フィロポディアはスパインの前駆体であるとの考えを提唱した(Ziv and Smith, 1996; Smith, 1999; Jontes
and Smith, 2000)。我々は,この「フィロポディアがスパインの前駆体である」という立場をとるが,前者二つの説を否定するデーターをもっているわけではない。なぜ我々がこの立場をとるのかについては後述する。
ところで,発達初期の段階で見られるフィロポディアのターンオーバーは非常に速く,大半はスパインにならずに消失する。そのため,フィロポディアはスパインの前駆体ではないとの立場をとるグループもある(Parnass et al., 2000; Portera-Cailliau
et al., 2003)。したがって,フィロポディアを形態から見てすべて同等と考えてはいけないのかもしれない。すなわち,発達初期の激しい動態を示すフィロポディアと,発達後期のスパイン前駆体フィロポディアとでは,形態は似ていても内在する状態が異なっていることが考えられる。
2.スパイン前駆体フィロポディアの分子マーカー
本当に状態の異なるフィロポディアが存在するのであろうか。もしも,スパインに移行するフィロポディアを見分ける分子マーカーがあれば,フィロポディアがスパイン前駆体であることを立証できる。
PSD-95は,スパイン前駆体を見分ける有力な分子マーカー候補である。Okabeらは,培養海馬神経細胞の樹状突起の形態変化とPSD-95クラスター出現をタイムラプスイメージングにより同時に追跡した(Okabe
et al., 2001)。樹状突起幹にもともと見られたPSD-95クラスターはスパイン形成には寄与せずに消失していく一方で,フィロポディア様の細い小突起の一部にはPSD-95の集積が起こることを示した。培養海馬スライスを用いた研究でも,PSD-95のあるスパインの前駆体がフィロポディアであることが示された
(Marrs et al., 2001)。また,PSD-95クラスターの存在する小突起は安定であることがわかっている(Prange and Murphy, 2001)。もしも,PSD-95が集まらないフィロポディアが消失するのであれば,PSD-95の集積の有無がフィロポディアの運命を決定すると言える。
では,スパインアクチン細胞骨格は,スパイン前駆体フィロポディアを見分けるマーカーになるであろうか。我々は最近,培養後14日の海馬神経細胞には,ドレブリンとアクチン繊維が瀰漫性に存在するタイプのフィロポディア(図1—B)以外に,ドレブリンがクラスターとして凝集しているフィロポディアが現れることを見いだした。フィロポディア内のドレブリンクラスター部位には,アクチン繊維が濃染しスパインアクチン細胞骨格の特徴を示しており,またシナプス小胞蛋白シナプシンIの集積部位が隣接していた。シナプスを持つフィロポディアにドレブリンクラスターが見られる確率はほぼ90%で,スパインに見られる頻度と同等であったが,PSD-95クラスターが見られる確率はそれより低く,約60%にとどまった。このことは,これらのフィロポディア内ではドレブリンクラスターの出現のほうがPSD-95クラスターの出現よりも早いことを示している。また,ドレブリンAアイソフォームの発現を抑制すると,ドレブリンクラスターが出現しなくなり,PSD-95もフィロポディアには集積せず,スパイン形成も阻害された(Takahashi
et al., 2003)。以上のことから,スパイン形成・成熟が進行するには,まずスパインアクチン細胞骨格がフィロポディア内に用意される必要があることがわかる。PSD-95などのアンカリングタンパクによりアクチン繊維が集められてスパインアクチン細胞骨格ができあがるという仮説では,この実験結果を説明出来ない。
これまで報告されたシナプス後部のタンパク集積のなかでは,ドレブリンのフィロポディア内集積が最も早い時期に起こることになる。Friedmanら(Friedman
et al., 2000)は,シナプス形成・成熟が,どのような段階を踏んで,どのような時間経過をたどるのかを調べた。シナプス形成は,軸索と樹状突起の接触から開始して,10−25分後にはシナプス前部にBassoonが集積し,20−35分後には機能的シナプス前アクティブゾーンの形成が起こり,遅れてシナプス小胞が集積する。そして60−70分後に,シナプス後部でPSD-95の集積が始まり,最後にグルタミン酸受容体がシナプス後膜に集積するという。シナプス後部の発達は,どうやら軸索との接触がその開始信号であるらしい。
しかし生後発達の初期には,フィロポディアと軸索の接触は必ずしもスパイン形成に至らないと考えられる。なぜなら,ドレブリンのクラスター形成はドレブリンAアイソフォームの発現に依存しているので,ドレブリンAアイソフォームが発現する以前(ラット生後1週齢以前)であれば,もしもフィロポディアが軸索に接触してもドレブリン結合型アクチン細胞骨格の出現は起こらないと考えられるからである。発達が進みドレブリンAが発現する時期に達すると,フィロポディアが軸索に接触することが引き金となり,ドレブリンがフィロポディア内でアクチンともにクラスターを形成し,それによりPSD-95がフィロポディア内に集積し,最終的にグルタミン酸受容体を集積させてスパインが完成すると思われる(図2;概念図)。ただし今のところ,フィロポディアとシナプス前部との接触からドレブリンクラスター形成までにかかる時間や,それに至るシグナルカスケードは不明である。ドレブリンクラスター形成を促すシグナルについては,接着因子やシナプス前部と後部の分化誘導因子などの研究から手がかりが得られるかもしれない。
最後に
今回、発達過程におけるスパイン形成のメカニズムにおいて,ドレブリンによるスパインアクチン細胞骨格の形成が主導的立場にあることを中心にまとめてみた。ドレブリン結合型アクチンの形成過程から、スパインはフィロポディアから形成されることや、ドレブリン結合型アクチンの集積がPSD分子のシナプス集積を制御していることが明らかとなった。また、最近の研究から、学習・記憶によりもたらされたシナプスの可塑的変化を長期的に保持するために,スパイン形態の可塑的変化が起こることが明らかになった。従って、ドレブリンは、シナプス可塑性に伴うスパイン新生にも同様に主導的立場を果たしている可能性が出てきた。
文献
1) Shepherd,
G.M.: J Neurophysiol,
75,2197-210 (1996)
2) Leuner, B., Falduto, J., and Shors, T.J.: J
Neurosci, 23,659-65 (2003)
3) Harris,
K.M., Jensen, F.E., and Tsao, B.: J Neurosci, 12,2685-705
(1992)
4) Boyer,
C., Schikorski, T., and Stevens, C.F.: J Neurosci, 18,5294-300 (1998)
5) Harris,
K.M. and Stevens, J.K.: J Neurosci, 9,2982-97 (1989)
6) Volfovsky, N., Parnas, H., Segal,
M., and Korkotian, E.: J Neurophysiol, 82,450-62 (1999)
7) Takumi,
Y., Ramirez-Leon, V., Laake, P., Rinvik,
E., and Ottersen, O.P.: Nat Neurosci, 2,618-24
(1999)
8) Matsuzaki, M., Ellis-Davies, G.C., Nemoto,
T., Miyashita, Y., Iino, M., and Kasai, H.:
Nat Neurosci, 4,1086-92 (2001)
9) Kasai,
H., Matsuzaki, M., Noguchi, J., Yasumatsu,
N., and Nakahara, H.: Trends Neurosci,
26,360-8 (2003)
10) Gray, E.: J. Anat. (Lond.),
93,420-433 (1959)
11) Nusser, Z., Mulvihill, E., Streit, P., and Somogyi, P.: Neuroscience, 61,421-7 (1994)
12) LeBeux, Y.J. and Willemot, J.: Cell Tissue Res,
160,1-36 (1975)
13) Matus, A.,
Ackermann, M., Pehling, G., Byers, H.R., and
Fujiwara, K.: Proc Natl Acad Sci U S A,
79,7590-4 (1982)
14) Fifkova, E. and Delay, R.J.: J Cell Biol, 95,345-50
(1982)
15) Fifkova, E.: Brain Res, 356,187-215
(1985)
16) Kennedy,
M.B.: Trends Neurosci, 20,264-8 (1997)
17) Ziff, E.B.: Neuron, 19,1163-74 (1997)
18) Sheng, M.
and Sala, C.: Annu
Rev Neurosci, 24,1-29 (2001)
19) Dong, H.,
O'Brien, R.J., Fung, E.T., Lanahan,
A.A., Worley, P.F., and Huganir, R.L.: Nature, 386,279-84 (1997)
20) Shirao, T.,
Kojima, N., Kato, Y., and Obata, K.:
Brain Res, 464,71-4
(1988)
21) Kojima, N.,
Kato, Y., Shirao, T., and Obata, K.:
Brain Res, 464,207-15
(1988)
22) Hayashi, K.,
Ishikawa, R., Ye, L.H., He, X.L., Takata, K., Kohama, K., and Shirao, T.: J Neurosci, 16,7161-70 (1996)
23) Shirao, T.,
Hayashi, K., Ishikawa, R., Isa, K., Asada, H., Ikeda, K., and Uyemura, K.: Exp Cell Res,
215,145-53 (1994)
24) Wyszynski, M., Lin, J., Rao, A.,
Nigh, E., Beggs, A.H., Craig, A.M., and Sheng, M.: Nature, 385,439-42 (1997)
25) Satoh, A.,
Nakanishi, H., Obaishi, H., Wada, M., Takahashi, K.,
Satoh, K., Hirao, K., Nishioka,
H., Hata, Y., Mizoguchi, A., and Takai,
Y.: J Biol Chem, 273,3470-5 (1998)
26) Deller, T., Merten, T., Roth,
S.U., Mundel, P., and Frotscher,
M.: J Comp Neurol, 418,164-81 (2000)
27) Rao, A. and Craig, A.M.:
Hippocampus, 10,527-41 (2000)
28) Fiala,
J.C., Feinberg, M., Popov, V., and Harris, K.M.: J Neurosci, 18,8900-11 (1998)
29) Harris,
K.M.: Curr Opin Neurobiol, 9,343-8 (1999)
30) Rao, A., Kim, E., Sheng, M., and Craig, A.M.: J Neurosci, 18,1217-29 (1998)
31) Ziv, N.E. and Smith, S.J.: Neuron, 17,91-102
(1996)
32) Smith,
S.J.: Science, 283,1860-1 (1999)
33) Jontes, J.D. and Smith, S.J.: Neuron, 27,11-4 (2000)
34) Parnass, Z., Tashiro, A., and Yuste, R.:
Hippocampus, 10,561-8 (2000)
35) Portera-Cailliau, C., Pan, D.T., and Yuste,
R.: J Neurosci, 23,7129-42 (2003)
36) Okabe, S.,
Miwa, A., and Okado, H.: J Neurosci, 21,6105-14
(2001)
37) Marrs, G.S., Green, S.H., and Dailey, M.E.: Nat Neurosci, 4,1006-13 (2001)
38) Prange, O. and Murphy, T.H.: J Neurosci, 21,9325-33
(2001)
39) Takahashi,
H., Sekino, Y., Tanaka, S., Mizui, T., Kishi, S., and Shirao, T.: J Neurosci, 23,6586-95 (2003)
40) Friedman,
H.V., Bresler, T., Garner, C.C., and Ziv, N.E.: Neuron,
27,57-69 (2000)
41) Qin, L., Marrs, G.S., McKim, R., and Dailey, M.E.: J Comp Neurol, 440,284-98
(2001)
42) Okabe, S.: Anat Sci Int, 77,203-10
(2002)
43) Garner,
C.C., Zhai, R.G., Gundelfinger,
E.D., and Ziv, N.E.: Trends Neurosci, 25,243-51
(2002)
44) Desmond,
N.L. and Levy, W.B.: J Comp Neurol, 253,476-82 (1986)
45) Toni, N., Buchs, P.A., Nikonenko, I., Povilaitite, P., Parisi, L., and
Muller, D.: J Neurosci, 21,6245-51 (2001)
46) Desmond,
N.L. and Levy, W.B.: J Comp Neurol, 253,466-75 (1986)
47) Toni, N., Buchs, P.A., Nikonenko, I., Bron, C.R., and Muller, D.: Nature, 402,421-5
(1999)
48) Fukazawa,
Y., Saitoh, Y., Ozawa, F., Ohta,
Y., Mizuno, K., and Inokuchi, K.: Neuron, 38,447-60
(2003)
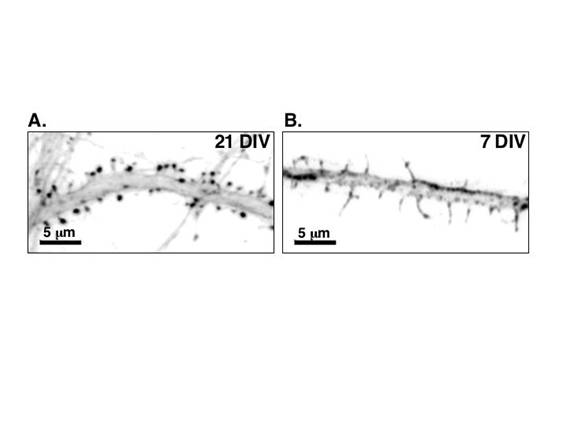
図1 低密度培養ラット海馬神経細胞樹状突起の抗ドレブリン抗体による免疫染色像
A: 培養21日後(21DIV)には,樹状突起スパインが完成している。スパイン頭部にドレブリンが集積している(本文II-2参照)。
B: 培養7日(7DIV)には,樹状突起に細長いフィロポディアが見られるのみである。ドレブリンは樹状突起の膜直下とフィロポディア内にびまん性に分布している(本文II-2参照)。
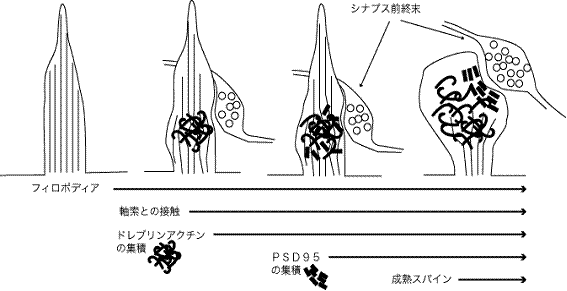
図2 樹状突起フィロポディアからスパインが完成するまでの分子過程
スパイン形成時におけるドレブリンアクチンとPSD-95のクラスターの発現時期の前後関係に着目してほしい。発達初期のフィロポディア内には非神経細胞と同様なアクチン細胞骨格が存在していると思われる。神経細胞に特異的なアクチン結合タンパクドレブリンAが発現する時期になると,フィロポディアと軸索の接触によりドレブリン結合型アクチン繊維がフィロポディア内に集積し始める。それにやや遅れてPSD-95の集積が開始する。最終的に成熟したスパイン内の階層的微細構造(PSDとスパインアクチ細胞骨格)が完成する。
Boyer C, Schikorski T,
Stevens CF (1998) Comparison of hippocampal dendritic spines in culture and in
brain. J Neurosci 18:5294-5300.
Desmond NL, Levy WB
(1986a) Changes in the postsynaptic density with long-term potentiation
in the dentate gyrus. J Comp Neurol 253:476-482.
Desmond NL, Levy WB
(1986b) Changes in the numerical density of synaptic contacts with long-term potentiation in the hippocampal dentate gyrus. J Comp
Neurol 253:466-475.
Fiala JC, Feinberg M, Popov V, Harris KM (1998) Synaptogenesis via dendritic
filopodia in developing hippocampal area CA1. J Neurosci 18:8900-8911.
Fifkova E (1985) Actin in the
nervous system. Brain Res 356:187-215.
Fifkova E, Delay RJ (1982)
Cytoplasmic actin in neuronal processes as a possible mediator of synaptic
plasticity. J Cell Biol 95:345-350.
Friedman HV, Bresler T, Garner CC, Ziv NE
(2000) Assembly of new individual excitatory synapses: time course and temporal
order of synaptic molecule recruitment. Neuron 27:57-69.
Fukazawa Y, Saitoh Y, Ozawa F, Ohta Y, Mizuno
K, Inokuchi K (2003) Hippocampal LTP is accompanied
by enhanced F-actin content within the dendritic spine that is essential for
late LTP maintenance in vivo. Neuron 38:447-460.
Harris KM (1999)
Structure, development, and plasticity of dendritic spines. Curr Opin Neurobiol
9:343-348.
Harris KM, Stevens JK
(1989) Dendritic spines of CA 1 pyramidal cells in the rat hippocampus: serial
electron microscopy with reference to their biophysical characteristics. J
Neurosci 9:2982-2997.
Harris KM, Jensen FE, Tsao B (1992) Three-dimensional structure of dendritic
spines and synapses in rat hippocampus (CA1) at postnatal day 15 and adult
ages: implications for the maturation of synaptic physiology and long-term potentiation. J Neurosci 12:2685-2705.
Hayashi K, Ishikawa R,
Ye LH, He XL, Takata K, Kohama K, Shirao T (1996) Modulatory
role of drebrin on the cytoskeleton within dendritic spines in the rat cerebral
cortex. J Neurosci 16:7161-7170.
Jontes JD, Smith SJ (2000)
Filopodia, spines, and the generation of synaptic diversity. Neuron 27:11-14.
Kennedy MB (1997) The
postsynaptic density at glutamatergic synapses. Trends Neurosci 20:264-268.
Kojima N, Kato Y,
Shirao T, Obata K (1988) Nucleotide sequences of two embryonic drebrins,
developmentally regulated brain proteins, and developmental change in their
mRNAs. Brain Res 464:207-215.
LeBeux YJ, Willemot J (1975) An ultrastructural study of the
microfilaments in rat brain by means of heavy meromyosin
labeling. I. The perikaryon, the dendrites and the
axon. Cell Tissue Res 160:1-36.
Leuner B, Falduto
J, Shors TJ (2003) Associative memory formation
increases the observation of dendritic spines in the hippocampus. J Neurosci
23:659-665.
Marrs GS, Green SH, Dailey
ME (2001) Rapid formation and remodeling of postsynaptic densities in
developing dendrites. Nat Neurosci 4:1006-1013.
Matsuzaki M, Ellis-Davies GC, Nemoto T, Miyashita Y, Iino M, Kasai H (2001) Dendritic
spine geometry is critical for AMPA receptor expression in hippocampal CA1
pyramidal neurons. Nat Neurosci 4:1086-1092.
Matus A, Ackermann M, Pehling G, Byers HR, Fujiwara K (1982) High actin
concentrations in brain dendritic spines and postsynaptic densities. Proc Natl
Acad Sci U S A 79:7590-7594.
Nusser Z, Mulvihill
E, Streit P, Somogyi P
(1994) Subsynaptic segregation of metabotropic
and ionotropic glutamate receptors as revealed by
immunogold localization. Neuroscience 61:421-427.
Okabe S, Miwa A, Okado H (2001) Spine formation and correlated assembly of
presynaptic and postsynaptic molecules. J Neurosci 21:6105-6114.
Parnass Z, Tashiro
A, Yuste R (2000) Analysis of spine morphological
plasticity in developing hippocampal pyramidal neurons. Hippocampus 10:561-568.
Portera-Cailliau C, Pan DT, Yuste R (2003) Activity-regulated dynamic behavior of early
dendritic protrusions: evidence for different types of dendritic filopodia. J
Neurosci 23:7129-7142.
Prange O, Murphy TH (2001)
Modular transport of postsynaptic density-95 clusters and association with
stable spine precursors during early development of cortical neurons. J
Neurosci 21:9325-9333.
Rao A, Craig AM (2000)
Signaling between the actin cytoskeleton and the postsynaptic density of
dendritic spines. Hippocampus 10:527-541.
Rao A, Kim E, Sheng M,
Craig AM (1998) Heterogeneity in the molecular composition of excitatory
postsynaptic sites during development of hippocampal neurons in culture. J
Neurosci 18:1217-1229.
Sheng M, Sala C (2001)
PDZ domains and the organization of supramolecular
complexes. Annu Rev Neurosci 24:1-29.
Shepherd GM (1996) The
dendritic spine: a multifunctional integrative unit. J Neurophysiol
75:2197-2210.
Shirao T, Kojima N,
Kato Y, Obata K (1988) Molecular cloning of a cDNA for the developmentally
regulated brain protein, drebrin. Brain Res
464:71-74.
Shirao T, Hayashi K,
Ishikawa R, Isa K, Asada H, Ikeda K, Uyemura K (1994) Formation of thick,
curving bundles of actin by drebrin A expressed in fibroblasts. Exp Cell Res 215:145-153.
Smith SJ (1999)
Dissecting dendrite dynamics. Science 283:1860-1861.
Takahashi H, Sekino Y,
Tanaka S, Mizui T, Kishi S, Shirao T (2003) Drebrin-dependent actin clustering
in dendritic filopodia governs synaptic targeting of postsynaptic density-95
and dendritic spine morphogenesis. J Neurosci 23:6586-6595.
Takumi Y, Ramirez-Leon
V, Laake P, Rinvik E, Ottersen OP (1999) Different modes of expression of AMPA
and NMDA receptors in hippocampal synapses. Nat Neurosci 2:618-624.
Toni N, Buchs PA, Nikonenko I, Bron CR, Muller D (1999) LTP promotes formation of multiple
spine synapses between a single axon terminal and a dendrite. Nature
402:421-425.
Toni N, Buchs PA, Nikonenko I, Povilaitite P, Parisi L, Muller D
(2001) Remodeling of synaptic membranes after induction of long-term potentiation. J Neurosci 21:6245-6251.
Volfovsky N, Parnas
H, Segal M, Korkotian E (1999) Geometry of dendritic
spines affects calcium dynamics in hippocampal neurons: theory and experiments.
J Neurophysiol 82:450-462.
Ziff EB (1997)
Enlightening the postsynaptic density. Neuron 19:1163-1174.
Ziv NE, Smith SJ (1996)
Evidence for a role of dendritic filopodia in synaptogenesis and spine
formation. Neuron 17:91-102.